



1. ņä£ ļĪĀ
ņĮöļĪ£ļéś ļ░öņØ┤ļ¤¼ņŖżļŖö ļŗżņ¢æĒĢ£ ļÅÖļ¼╝ņØä Ļ░ÉņŚ╝ņŗ£ĒéżĻ│Ā ņØĖĻ░äņŚÉĻ▓ī Ļ▓Įņ”ØņŚÉņä£ ņżæņ”ØņØś ĒśĖĒØĪĻĖ░ Ļ░ÉņŚ╝ņ¦łĒÖśņØä ņ£Āļ░£ĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż[1]. 2002ļģäĻ│╝ 2012ļģäņŚÉ ĒØöĒĢ£ Ļ░ÉņŚ╝ļ│æņØä ņØ╝ņ£╝ĒéżļŖö Ļ│Āļ│æņøÉņä▒ ņĮöļĪ£ļéśļ░öņØ┤ļ¤¼ņŖżņØĖ ņżæņ”ØĻĖēņä▒ĒśĖĒØĪĻĖ░ņ”ØĒøäĻĄ░(SARS-CoV-2)Ļ│╝ ņżæļÅÖĒśĖĒØĪĻĖ░ņ”ØĒøäĻĄ░(MERS-CoV-2)ņØ┤ ņé¼ļ×īņŚÉĻ▓ī ņ╣śļ¬ģņĀüņØĖ ĒśĖĒØĪĻĖ░ ņ¦łĒÖśņØä ņØ╝ņ£╝ņ╝£ ņŗĀņóģ ņĮöļĪ£ļéśļ░öņØ┤ļ¤¼ņŖżļź╝ 21ņäĖĻĖ░ņØś ņāłļĪ£ņÜ┤ Ļ│Ąņżæļ│┤Ļ▒┤ ļ¼ĖņĀ£ļĪ£ ļČĆĻ░üļÉśņŚłļŗż[1]. 2019ļģä ļ¦É, SARS-CoV-2ļĪ£ ļ¬ģļ¬ģļÉ£ ņŗĀņóģ ņĮöļĪ£ļéśļ░öņØ┤ļ¤¼ņŖżĻ░Ć ņżæĻĄŁ ņÜ░ĒĢ£ņŗ£ņŚÉņä£ ņČ£ĒśäĒĢśņŚ¼ ņØ┤ļĪĆņĀüņØĖ ļ░öņØ┤ļ¤¼ņŖżņä▒ ĒÅÉļĀ┤ņØ┤ ļ░£ņāØĒĢśņśĆļŗż[1,2]. ņĮöļĪ£ļéśļ░öņØ┤ļ¤¼ņŖż 2019(COVID-19)ļĪ£ ņĢīļĀżņ¦ä ņŗĀņóģ ņĮöļĪ£ļéśļ░öņØ┤ļ¤¼ņŖż ņ¦łļ│æņØĆ ļåÆņØĆ Ļ░ÉņŚ╝ļĀźņ£╝ļĪ£ ņØĖĒĢ┤ ņĀä ņäĖĻ│äņĀüņ£╝ļĪ£ ļ╣Āļź┤Ļ▓ī ĒŹ╝ņĪīļŗż[3].
ņ×ÉņŚ░ņé┤ĒĢ┤ņäĖĒż(NKņäĖĒż)ļŖö ļ│Ėļל ļ░öņØ┤ļ¤¼ņŖż ļśÉļŖö ņóģņ¢æņäĖĒżņŚÉ ņ×ÉņŚ░ņĀüņØĖ ņäĖĒżļÅģņä▒ņØä Ļ░Ćņ¦ĆļŖö Ēü░ Ļ│╝ļ”Į ļ”╝ĒöäĻĄ¼ļĪ£ ņäżļ¬ģļÉśņŚłļŗż[4,5]. NKņäĖĒżļŖö ļéśņżæņŚÉ ļ╣äņĀĢņāü ņäĖĒżņŚÉ ļīĆĒĢ£ ņäĖĒżļÅģņä▒Ļ│╝ ņé¼ņØ┤ĒåĀņ╣┤ņØĖ ņāØņä▒ņØä ĒåĄĒĢ£ ļŗżļźĖ ļ®┤ņŚŁ ņäĖĒżņØś ĒÖ£ņä▒ņØä ņ”ØĻ░Ćņŗ£ĒéżļŖö ļŖźļĀźņØä ļ¬©ļæÉ Ļ░Ćņ¦ä ļ│äļÅäņØś ļ”╝ĒöäĻĄ¼ Ļ│äĒåĄņ£╝ļĪ£ ņØĖņŗØļÉśņŚłļŗż[4,6]. ņ¦äĒÖö Ļ│╝ņĀĢņŚÉņä£ Ēæ£ņĀü ņäĖĒżņŚÉ ļīĆĒĢ£ ņäĖĒżļÅģņä▒ņØä ĒÜŹļōØĒĢśļŖö Ļ▓āņØĆ ņäĖĒż ņÜ®ĒĢ┤ Ļ│╝ņĀĢņØä ņĀ£ņ¢┤ĒĢśļ®┤ņä£ ņĀĢņāü ņĪ░ņ¦ü ņåÉņāüņØä ļ░®ņ¦ĆĒĢśļŖö Ļ│ĀļÅäļĪ£ ņĀĢĻĄÉĒĢśĻ│Ā Ļ░ĢļĀźĒĢ£ ļ®öņ╗żļŗłņ”śņØś Ļ░£ļ░£Ļ│╝ Ļ┤ĆļĀ©ņØ┤ ņ׳ļŗż[4,7]. ņØ┤ņÖĆ Ļ┤ĆļĀ©ĒĢśņŚ¼ NKņäĖĒżĻ░Ć Ēæ£ņĀüņäĖĒżļź╝ ļŗżļźĖ Ļ▒┤Ļ░ĢĒĢ£ ŌĆśņ×ÉĻĖ░(self)ŌĆÖ ņäĖĒżņÖĆ ĻĄ¼ļ│äĒĢĀ ņłś ņ׳ļÅäļĪØ ĒĢśļŖö ļ®öņ╗żļŗłņ”śņØä ĒÖĢņØĖĒĢśļŖöļŹ░ ņāüļŗ╣ĒĢ£ ņ¦äņĀäņØ┤ ņ׳ņŚłļŗż[4,8]. NKņäĖĒżņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼ļŖö NKņäĖĒż Ēæ£ļ®┤ņŚÉ ņĪ┤ņ×¼ĒĢśļŖö ļŗżņ¢æĒĢ£ ĒÖ£ņä▒ĒÖö ļ░Å ņ¢ĄņĀ£ ņłśņÜ®ņ▓┤ņÖĆ Ļ┤ĆļĀ©ņØ┤ ņ׳ņ£╝ļ®░, ņØ┤ļ¤¼ĒĢ£ ņłśņÜ®ņ▓┤ņŚÉ ĒĢ┤ļŗ╣ĒĢśļŖö ļ”¼Ļ░äļō£ņØś Ļ▓░ĒĢ®ņØ┤ NKņäĖĒżņØś ĒÖ£ņä▒ņØä ņĪ░ņĀłĒĢśļŖö ĒĢ£ ļ®öņ╗żļŗłņ”śņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż[4,9]. ļö░ļØ╝ņä£ ņØ┤ņøā ņäĖĒżņÖĆņØś ņāüĒśĖ ņ×æņÜ®ņŚÉņä£ ĻĖĖĒĢŁĻ▓ĮļĪ£ņØś ĒåĄĒĢ®ņØĆ NKņäĖĒżņØś ĒÖ£ņä▒ĒÖöļź╝ ņĪ░ņĀłĒĢśļŖö ļÅÖņĀü ĒÅēĒśĢņØä ņ¦Ćļ░░ĒĢśĻ│Ā, NKņäĖĒżļŖö ĒÖ£ņä▒ĒÖöļÉśņ¢┤ Ēæ£ņĀüņäĖĒżņØś ņé¼ļ®Ė ņŚ¼ļČĆļź╝ Ļ▓░ņĀĢ ĒĢ£ļŗż[4].
NKņäĖĒż ĻĖ░ļŖźņØĆ ņŚ¼ļ¤¼ ņĢöĒśĖĒÖöļÉ£ ņłśņÜ®ņ▓┤ņŚÉ ņØśĒĢ┤ ņĪ░ņĀłļÉśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż[10,11]. ļ░öņØ┤ļ¤¼ņŖż Ļ░ÉņŚ╝ ņŗ£, ņłÖņŻ╝ ņäĖĒżļŖö Ļ░ÉņŚ╝ņŚÉ ņØśĒĢ┤ ņ£ĀļÅäļÉ£ ņ×Éņ▓┤ ņĢöĒśĖĒÖö ļČäņ×ÉņØś ņāüĒ¢ź ņĪ░ņĀł ļ░Å/ļśÉļŖö ņäĖĒżļÅģņä▒ ņłśņÜ®ņ▓┤(ex, NKp30, NKp44 ļ░Å NKp46), CĒśĢ lectin ņ£Āņé¼ ņłśņÜ®ņ▓┤(ex, NKG2D ļ░Å NKp80) ļ░Å Ļ│ĄļÅÖĒÖ£ņä▒ĒÖö ņłśņÜ®ņ▓┤(ex, DNAM-1 ļ░Å CD2)ņÖĆ Ļ░ÖņØĆ ĒÖ£ņä▒ĒÖö NKņäĖĒż ņłśņÜ®ņ▓┤ņŚÉ Ļ▓░ĒĢ®ĒĢ┤ ņłśļ░śļÉśļŖö ņäĖĒż ņŖżĒŖĖļĀłņŖż ļ░śņØæņØä ĒżĒĢ©ĒĢśļŖö ļŗżņ¢æĒĢ£ ļ®öņ╗żļŗłņ”śņØä ĒåĄĒĢ┤ NKņäĖĒż ļ¦żĻ░£ ņØĖņŗØņŚÉ ļ»╝Ļ░ÉĒĢ┤ņ¦ł ņłś ņ׳ļŗż[10]. ļśÉĒĢ£ ņ¢ĄņĀ£ ņłśņÜ®ņ▓┤ņŚÉ ļīĆĒĢ£ MHC class I ligandņØś ĒĢśĒ¢źņĪ░ņĀłņØĆ Ēæ£ņĀüņäĖĒż ļ»╝Ļ░Éņä▒ ņ”ØĻ░ĆņŚÉ ĻĖ░ņŚ¼ĒĢ£ļŗż[10]. NKņäĖĒżļŖö ļśÉĒĢ£ CD16 ļ¦żĻ░£ ĒĢŁņ▓┤ ņØśņĪ┤ņä▒ ņäĖĒżļÅģņä▒(ADCC)ņØä ĒåĄĒĢ┤ ļ░öņØ┤ļ¤¼ņŖżņŚÉ Ļ░ÉņŚ╝ļÉ£ ņäĖĒżļź╝ ņĀ£Ļ▒░ĒĢĀ ņłśļÅä ņ׳ļŗż[10]. ļ¦łņ¦Ćļ¦ēņ£╝ļĪ£, NK ņäĖĒżņØś ĒÖ£ņä▒ņØĆ ņØĖĒä░ļŻ©Ēé© IL-2, IL-12, IL-15, IL-18 ļ░Å type I ņØĖĒä░ĒÄśļĪĀņØä ĒżĒĢ©ĒĢśļŖö ņé¼ņØ┤ĒåĀņ╣┤ņØĖļōżņŚÉ ņØśĒĢ┤ ņĪ░ņĀłļÉ£ļŗż[10]. ĒĢśņ¦Ćļ¦ī ņØ┤ļ¤¼ĒĢ£ ņĪ░ņĀłņØĆ ļ░öņØ┤ļ¤¼ņŖż Ļ░ÉņŚ╝ ņäĖĒż ļśÉļŖö ĒÖ£ņä▒ĒÖöļÉ£ ĒĢŁņøÉ ņĀ£ņŗ£ ņäĖĒżņŚÉ ņØśĒĢ┤ ņāØņä▒ļÉĀ ņłś ņ׳ļŖö ņé¼ņØ┤ĒåĀņ╣┤ņØĖņŚÉ ĻĄŁĒĢ£ļÉśņ¦Ć ņĢŖņØä ņłś ņ׳ļŗż[10,12]. ņØ┤ļ¤¼ĒĢ£ ļ¦ÄņØĆ ņé¼ņØ┤ĒåĀņ╣┤ņØĖņØĆ ļŗ©ļÅģņ£╝ļĪ£ ļśÉļŖö ņĪ░ĒĢ®ĒĢśņŚ¼ NKņäĖĒż ņāØņĪ┤, ņ”ØņŗØ, ņäĖĒżļÅģņä▒ ļ░Å ņØĖĒä░ĒÄśļĪĀ Ļ░Éļ¦ł(IFN-╬│) ņāØņé░ņØä ĒżĒĢ©ĒĢśļŖö ņé¼ņØ┤ĒåĀņ╣┤ņØĖ ņāØņä▒ņØä ņ┤ēņ¦äĒĢ£ļŗż[13]. ņØ┤ļ¤¼ĒĢ£ ļ®öņ╗żļŗłņ”śņØä ĒåĄĒĢ┤ NKņäĖĒżļŖö ļ░öņØ┤ļ¤¼ņŖż Ļ░ÉņŚ╝ ņäĖĒżļź╝ Ļ░Éņ¦ĆĒĢśĻ│Ā ņ”ēĻ░üņĀüņ£╝ļĪ£ ņĀ£Ļ▒░ĒĢĀ ņłś ņ׳ļŗż[10].
NKņäĖĒżņØś ex-vivo ļ░░ņ¢æņØä ņ£äĒĢ┤ ļ”╝ĒöäĻĄ¼ņØś ļČäļ”¼ļŖö ņŻ╝ļĪ£ Ficoll-Hypaque ļ░ĆļÅä ĻĄ¼ļ░░ ņøÉņŗ¼ļČäļ”¼ļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ļ”╝ĒöäĻĄ¼ļź╝ ļČäļ”¼ĒĢśĻ│Ā[14], ļČäļ”¼ļÉ£ ņäĖĒżņŚÉņä£ IL-2ņÖĆ Ļ░ÖņØĆ ņäĖĒż ņ”ØņŗØņŚÉ Ļ┤ĆņŚ¼ĒĢśļŖö ņé¼ņØ┤ĒåĀņ╣┤ņØĖĻ│╝ ĒĢ©Ļ╗ś ņ¦Ćņ¦ĆņäĖĒż(feeder cell)ņÖĆ Ļ│ĄļÅÖļ░░ņ¢æ ĒĢśļŖö ļ░®ļ▓ĢņØ┤ ņé¼ņÜ®ļÉ£ļŗż[10,15,16]. ņ×äņāüņĀüņ£╝ļĪ£ ņé¼ņÜ®ĒĢśĻĖ░ ņ£äĒĢ£ NKņäĖĒż ļ░░ņ¢æņØĆ Ļ│Āņł£ļÅä, ņĀĆļČłņł£ļÅäļź╝ ļ│┤ņןĒĢ┤ņĢ╝ ĒĢśņ¦Ćļ¦ī ņ¦Ćņ¦ĆņäĖĒżņÖĆņØś Ļ│ĄļÅÖ ļ░░ņ¢æ ļō▒ņØś Ļ│╝ņĀĢņŚÉņä£ ĒÜ©Ļ│╝ņĀüņØĖ ņé¼ņØ┤ĒåĀņ╣┤ņØĖ ņäĀļ│ä ļ░Å ļČłņł£ļ¼╝ ņĀ£ņ¢┤Ļ░Ć ņ¢┤ļĀĄļŗżļŖö ņĀÉņØ┤ NKņäĖĒż ļ░░ņ¢æņØś ļé£Ļ┤ĆņØ┤ļŗż. ļö░ļØ╝ņä£ ņ£äņÖĆ Ļ░ÖņØĆ ņØ┤ņ£ĀļĪ£ NKņäĖĒżļź╝ ņØ┤ņÜ®ĒĢ£ ņäĖĒżņ╣śļŻīņĀ£ņØś Ļ░£ļ░£ņØ┤ ņ¢┤ļĀĄĻ│Ā, ļ░░ņ¢æ ļ░®ļ▓ĢņØś Ēæ£ņżĆĒÖöĻ░Ć ĒĢäņÜöĒĢ£ ņŗżņĀĢņØ┤ļŗż[17,18].
ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö ļ¦Éņ┤łĒśłņĢĪņŚÉņä£ ļČäļ”¼ļÉ£ ļ¦Éņ┤łĒśłņĢĪļŗ©ĒĢĄņäĖĒżļĪ£ļČĆĒä░ NKņäĖĒżļź╝ ļČäļ”¼ĒĢśĻ│Ā, ļ░░ņ¢æ ļ░®ļ▓ĢņØä Ēæ£ņżĆĒÖöĒĢśņŚ¼ ņ×äņāüņĀüņ£╝ļĪ£ ņĀüņÜ®(ņäĖĒżņłś: >1ņ¢Ą)ņØ┤ Ļ░ĆļŖźĒĢĀ[19] ņĀĢļÅäņØś ļīĆņÜ®ļ¤ēņ£╝ļĪ£ ļ░░ņ¢æņØ┤ Ļ░ĆļŖźĒĢ£ ĒĢŁļ░öņØ┤ļ¤¼ņŖż NKņäĖĒż ļ░░ņ¢æĻ│ĄņĀĢ Ēöīļ×½ĒÅ╝ņØä ĒÖĢļ”ĮĒĢśĻ│Ā, ļ░░ņ¢æļÉ£ NKņäĖĒżņØś SARS-CoV-2 ļ░öņØ┤ļ¤¼ņŖżņŚÉ ļīĆĒĢ£ ĒĢŁļ░öņØ┤ļ¤¼ņŖż ĒÜ©ļŖźņØä ĒÖĢņØĖĒĢśņŚ¼ ņ×äņāüņŚÉ ņĀüņÜ® Ļ░ĆļŖźĒĢ£ ĒĢŁļ░öņØ┤ļ¤¼ņŖż ņäĖĒżņ╣śļŻīņĀ£ Ēøäļ│┤ĻĄ░ Ļ░£ļ░£ņØś ĻĖ░ĒŗĆņØä ļ¦łļĀ©ĒĢśĻ│Āņ×É ĒĢśņśĆļŗż.
2. Material and methods
2.1 Materials
A549 ļ░Å VERO E6 ņäĖĒżņŻ╝ļŖö American Type Culture Collection(ATCC, Manassas, VA, USA)ņŚÉņä£ ĻĄ¼ņ×ģĒĢśņśĆĻ│Ā Dulbecco's Modified Eagle Medium(DMEM) ļ░░ņ¢æļ░░ņ¦ĆļŖö Gibco(Billings, MT, USA)ņŚÉņä£ ĻĄ¼ņ×ģĒĢśņśĆļŗż. Fetal Bovine Serum(FBS)ņØĆ Hyclone(Logan, UT USA)ņŚÉņä£ ĻĄ¼ņ×ģĒĢśņśĆĻ│Ā, NKņäĖĒż ļ░░ņ¢æļ░░ņ¦ĆņØĖ KBM502ļŖö KojinBio(Sakado, Japan)ņŚÉņä£ ĻĄ¼ņ×ģĒĢśņśĆļŗż. EasySep TM Human NK Cell Isolation Kit, Human CD3 Positive Selection Kit II ļ░Å The Big EasySep TM magnetņØĆ STEMCELL Technologies (Seoul, Korea)ņŚÉņä£ ĻĄ¼ņ×ģĒĢśņśĆļŗż. Proteome Profiler Human XL Cytokine Array KitļŖö R&D Systems(Minneapolis, MN, USA)ņŚÉņä£ ĻĄ¼ņ×ģĒĢśņśĆļŗż. Poly-L-lysine solutionĻ│╝ Bovine Serum AlbuminņØĆ Sigma-Aldrich(Seoul, Korea)ņŚÉņä£ ĻĄ¼ņ×ģĒĢśņśĆļŗż. Paraformaldehyde(4 %)ļŖö T&I(Zhangjiakou, China)ņŚÉņä£, 0.4 % Trypan Blue StainņØĆ Thermo Fisher Scientific(Waltham, MA, USA)ņŚÉņä£ ĻĄ¼ņ×ģĒĢśņśĆļŗż. CD3, CD56, CD107a, NKG2D ļ░Å CD57 ĒĢŁņ▓┤ļŖö Invitrogen (Waltham, MA, USA)ņŚÉņä£ ĻĄ¼ņ×ģĒĢśņśĆļŗż. CD16, NKG2C, NKp30, NKp44, NKp46, NKp80, DNAM-1, 2B4, FAS-L ļ░Å TRAIL ĒĢŁņ▓┤ļŖö BioLegend(SanDiego, CA, USA)ņŚÉņä£ ĻĄ¼ņ×ģĒĢśņśĆļŗż. NKsol TMņØĆ IMMUNISBIO Co., Ltd(Incheon, Korea)ņŚÉņä£ ņĀ£Ļ│Ą ļ░øņĢśļŗż. SARS-CoV-2 delta variantļŖö ņ¦łļ│æĻ┤Ćļ”¼ļ│ĖļČĆ(Cheongju, Korea)ņŚÉņä£ ļ░░ĒżļÉśņ¢┤ biosafety level 3 ņŗżĒŚśņŗżņŚÉņä£ ļ│┤Ļ┤ĆĒĢśņśĆļŗż. ņØĖĻ░ä ļ¦Éņ┤ł ĒśłņĢĪ(PB) ņāśĒöīņØś Ļ▓Ćņé¼ļŖö Ļ░ĆĒå©ļ”ŁĻ┤ĆļÅÖļīĆĒĢÖĻĄÉ ĻĄŁņĀ£ņä▒ļ¬©ļ│æņøÉ ņ×äņāüņŚ░ĻĄ¼ņŗ¼ņØśņ£äņøÉĒÜī(IRB committee)ņØś ņŖ╣ņØĖņØä ļ░øņĢä ņłśĒ¢ēļÉśņŚłļŗż.
2.2 Methods
2.2.1 NK cell preparation
Ļ▒┤Ļ░ĢĒĢ£ ĻĖ░ņ”Øņ×ÉļĪ£ļČĆĒä░ ņ¢╗ņØĆ ļ¦Éņ┤ł ĒśłņĢĪ(PB)ņŚÉņä£ ļČäļ”¼ļÉ£ ļ¦Éņ┤ł ĒśłņĢĪ ļŗ©ĒĢĄ ņäĖĒż(PBMC)ļŖö Ficoll-Hypaque ļ░ĆļÅä ĻĄ¼ļ░░ ņøÉņŗ¼ļČäļ”¼ļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ļČäļ”¼ĒĢśņśĆļŗż. ļČäļ”¼ļÉ£ PBMC ļź╝ Ļ░üĻ░ü EasySepTM Human NK Cell Isolation Kit ļ░Å EasySep TM Human CD3 Positive Selection Kit IIļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ņĀ£ņĪ░ņŚģņ▓┤ņØś ņ¦Ćņ╣©ņŚÉ ļö░ļØ╝ NKņäĖĒż(NKi ņäĖĒż) ļ░Å CD3+ depletion ņäĖĒż(CD3-)ļź╝ ļČäļ”¼ĒĢśņśĆļŗż. ņÜöņĢĮĒĢśņŚ¼, Big Easy EasySep TM ņ×ÉņäØņØä ņé¼ņÜ®ĒĢśņŚ¼ ļČäļ”¼ļÉ£ PBļź╝ ĒĢ┤ļŗ╣ kitņØś selected cocktail concentrationsļĪ£ ļ░śņØæņŗ£ĒéżĻ│Ā RapidSpheres TMļĪ£ ņ▓śļ”¼ĒĢśņŚ¼ ņäĖĒżļź╝ ļČäļ”¼ĒĢśņśĆļŗż. ļČäļ”¼ĒĢ£ ņäĖĒżļź╝ ņĀ£ņĪ░ņŚģņ▓┤ņØś ņ¦Ćņ╣©ņŚÉ ļö░ļØ╝ KBM502 ļ░░ņ¦ĆņŚÉ ņ”ØņŗØ ļ░Å ĒÖ£ņä▒ĒÖö ņŗ£ņĢĮņØĖ NKsol TMņ£╝ļĪ£ ņ▓śļ”¼ĒĢśņŚ¼ ļ░░ņ¢æĒĢśņśĆļŗż. ļČäļ”¼ ļ░░ņ¢æļÉ£ NKi ļ░Å CD3-ņäĖĒż(20├Ś106 cells)ņŚÉ 0ņØ╝(D0)ļČĆĒä░ 6ņØ╝(D6)Ļ╣īņ¦Ć 3ņØ╝ļ¦łļŗż 20 ╬╝LņØś NKsol TMņØä ņ▓śļ”¼ĒĢśņŚ¼ ļ░░ņ¢æĒĢśņśĆļŗż.
2.2.2 Expansion kinetics assesment
Ļ░ü NKiņäĖĒżņÖĆ CD3-ņäĖĒżņØś ņ┤Ø ņäĖĒż ņłśļŖö Trypan Blue StainņØä ņé¼ņÜ®ĒĢśņŚ¼ 0ņØ╝(D0), 3ņØ╝(D3), 5ņØ╝(D5), 8ņØ╝(D8) ļ░Å 16ņØ╝(D16)ņŚÉ ņĖĪņĀĢļÉśņŚłļŗż. ņĀäņ▓┤ ņģĆ ņłśņØś ļ│ĆĒÖöļŖö Ļ░ü ĒżņØĖĒŖĖņØś ņäĖĒż ņłśļź╝ D0 ņäĖĒż ņłśļĪ£ ļéśļłäņ¢┤ fold expansionņ£╝ļĪ£ Ēæ£ĒśäĒĢśņśĆļŗż.
2.2.3 Flow cytometry
NKi ļ░Å CD3-ņäĖĒżļŖö CD3, CD56, CD16, CD107a, NKG2D, NKG2C, NKG2A, NKp30, NKp44, NKp46, NKp80, DNAM-1, 2B4, FAS-L, CD57 ļ░Å TRAIL ĒĢŁņ▓┤ļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ļČäņäØĒĢśņśĆļŗż. ņ£ĀņäĖĒż ļČäņäØņØĆ CytoFLEX (Beckman Coulter, Brea, CA)ļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ņłśĒ¢ēĒĢśņśĆļŗż. ļŹ░ņØ┤Ēä░ļŖö CytoExpert ņåīĒöäĒŖĖņø©ņ¢┤ļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ļČäņäØĒĢśņśĆļŗż.
2.2.4 Antiviral effect of natural killer cells using the TCID50 assay
NKņäĖĒżņØś ĒĢŁļ░öņØ┤ļ¤¼ņŖż ĒÜ©Ļ│╝ļź╝ ĒÖĢņØĖĒĢśĻĖ░ ņ£äĒĢ┤ VERO E6 ņäĖĒżļź╝ 100 % confluenceļĪ£ ļ░░ņ¢æĒĢśĻ│Ā 1├Ś104 PFU/mL ļĪ£ SARS-CoV-2 delta varientļź╝ N-p-Tosyl-L-phenylalanine chloromethyl ketone(TPCK)-trypsinĻ│╝ ĒĢ©Ļ╗ś ņ▓śļ”¼ĒĢśņŚ¼ 2ņŗ£Ļ░ä ļÅÖņĢł ļ░░ņ¢æĒĢśņśĆļŗż. Fresh mediumņ£╝ļĪ£ ĻĄÉņ▓┤ Ēøä, NKņäĖĒż Ēøäļ│┤ĻĄ░ņØś Ļ│Ąļ░░ņ¢æņŚÉ ļö░ļźĖ virus titer ļ│ĆĒÖöļź╝ ĒÖĢņØĖĒĢśĻĖ░ ņ£äĒĢ┤ VERO E6ņäĖĒż ļŗ©ļÅģ ĻĄ░(control)Ļ│╝ VERO E6ļź╝ Ēæ£ņĀüņäĖĒżļĪ£ ĒĢśļŖö ņØ┤ĒÄÖĒä░ņäĖĒżņØĖ NKņäĖĒż Ēøäļ│┤ĻĄ░ņØä 1:1, 1:5ņØś ņäĖĒż ņłś ļ╣äņ£©ļĪ£ Ļ│Ąļ░░ņ¢æ ĒĢśņśĆļŗż. Ļ│Ąļ░░ņ¢æņ£╝ļĪ£ ļČĆĒä░ ĒÜŹļōØĒĢ£ ļ░░ņ¢æņĢĪņØä 13,000 rpmņ£╝ļĪ£ 5ļČäĻ░ä ņøÉņŗ¼ļČäļ”¼ĒĢśņŚ¼ supernatantļź╝ ĒÜŹļōØĒĢśņŚ¼ TCID50 assayņŚÉ ņé¼ņÜ®ĒĢśņśĆļŗż. ņĢ×ņä£ ņ¢╗ņØĆ viral supernatantļŖö 96 well plate ņŚÉ 100 % confluencyļĪ£ ļ░░ņ¢æļÉ£ VERO E6ņŚÉ ņĀüņÜ®ĒĢśĻ│Ā, viral titerļŖö cytopathic effect(CPE)ļź╝ Ļ┤Ćņ░░ĒĢśņŚ¼ ļČäņäØĒĢśņśĆļŗż. ņĀäņ▓┤ ņŗżĒŚś Ļ│╝ņĀĢņØĆ biosafety level 3 laboratoryņŚÉņä£ ņłśĒ¢ēĒĢśņśĆļŗż.
2.2.5 in vivo experiment hACE transgenic (hACE-tg) mouse model
ļ¬©ļōĀ ļÅÖļ¼╝ņŗżĒŚśņØĆ ĻĄŁļ”ĮņŚ░ĻĄ¼ĒśæņØśĒÜī ņŗżĒŚśļÅÖļ¼╝ Ļ┤Ćļ”¼ ļ░Å ņé¼ņÜ®ņ¦Ćņ╣©(IACUC, Korea, ADD-IACUC-22-15)ņŚÉ ļö░ļØ╝ ņłśĒ¢ēĒĢśņśĆļŗż.
ļ¬©ļōĀ ļ¦łņÜ░ņŖżļŖö Jackson ņŚ░ĻĄ¼ņåīņŚÉņä£ ĻĄ¼ņ×ģĒĢśņŚ¼ 21Ōł╝23 ŌäāņØś ņś©ļÅäņŚÉņä£ 12ņŗ£Ļ░ä ļ░żļé« ņŻ╝ĻĖ░ļĪ£ ļ¼╝Ļ│╝ ņØīņŗØņŚÉ ņĀæĻĘ╝ĒĢĀ ņłś ņ׳ļŖö ABL3 ĒÖśĻ▓ĮņŚÉņä£ ņ£Āņ¦ĆļÉśņ¢┤ ņé¼ņ£ĪļÉśņŚłļŗż.
hACE-tg ļ¦łņÜ░ņŖż(B6.Cg-Tg(K18-ACE2)2Prlmn/J)ļŖö 1├Ś107 cells/headļĪ£ PBMC ļīĆņĪ░ĻĄ░, NKi, CD3-ņäĖĒżļź╝ ņŻ╝ņ×ģĒĢśĻ│Ā 1ņŗ£Ļ░ä Ēøä SARS-CoV-2 delta variant 1,700 PFU/headļź╝ intranasally(i.n.)ļĪ£ Ļ░ÉņŚ╝ņŗ£ņ╝░ļŗż. Weight changeņÖĆ survival ņØĆ 2ņŻ╝ ļÅÖņĢł ņĖĪņĀĢļÉśņŚłļŗż. ļ¦łņÜ░ņŖżņØś ņ┤Ø ļ¦łļ”¼ ņłśļŖö ĻĘĖļŻ╣ ļŗ╣ 9ļ¦łļ”¼ļź╝ ņé¼ņÜ®ĒĢśņśĆļŗż. ļ¬©ļōĀ ļ¦łņÜ░ņŖżļŖö ņĀĆņÜ®ļ¤ē SARS-CoV-2 Ļ░ÉņŚ╝ņŚÉņä£ ņāØņĪ┤ņ£©ņØ┤ ļé«ņØĆ ņłśņ╗ĘņØ┤ņŚłļŗż. ļ»ĖĻĄŁ ņłśņØśĒĢÖ ĒśæĒÜī ņ¦Ćņ╣©ņŚÉ ļö░ļØ╝ ļ¬©ļōĀ ņāØņĪ┤ ļÅÖļ¼╝ņØä 10 % CRR ņŚÉņä£ CO2ļĪ£ ņĢłļØĮņé¼ ĒĢśņśĆļŗż.
3. Results
3.1 Natural Killer cell sources were determined through isolation methods, and natural killer cell therapeutic candidates were identified through culture
NKņäĖĒż ļ░░ņ¢æņØä ņ£äĒĢ┤ Ficoll-Hypaque ļ░ĆļÅäĻĄ¼ļ░░ ņøÉņŗ¼ļČäļ”¼ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ Ļ▒┤Ļ░ĢĒĢ£ Ļ│ĄņŚ¼ņ×ÉņØś PBņŚÉņä£ ļ”╝ĒöäĻĄ¼ļź╝ ļČäļ”¼ĒĢśņśĆļŗż. ņØ┤Ēøä, ņČöĻ░ĆņĀüņ£╝ļĪ£ ņł£ņłś NKņäĖĒż Ēøäļ│┤ĻĄ░ņØä ņäĀļ│äĒĢśĻ│Ā ļŗżņ¢æĒĢ£ ņé¼ņØ┤ĒåĀņ╣┤ņØĖņØä ĒżĒĢ©ĒĢśļŖö ļ░░ņ¦ĆļĪ£ ĒÖ£ņä▒ ļ░Å/ļśÉļŖö ņ”ØņŗØ NKņäĖĒżļź╝ ņ¢╗ņ¢┤ ņĄ£ņóģ Ēøäļ│┤ĻĄ░ņØä ļ░░ņ¢æĒĢ£ļŗż. ņØ╝ļ░śņĀüņ£╝ļĪ£ ņĢīļĀżņ¦ä NKņäĖĒż ļ░░ņ¢æņØĆ feeder ņäĖĒżņØś Ļ│Ąļ░░ņ¢æņØ┤ļéś ļŗżļŗ©Ļ│äņØś ļČäļ│ä ļ░®ļ▓Ģ ļ│Ąņ×ĪĒĢ£ Ļ│ĄņĀĢņØä ņé¼ņÜ®ĒĢśĻ│Ā ņ׳ļŗż. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö Ļ░äļŗ©ĒĢ£ ļ░░ņ¢æ Ēöīļ×½ĒÅ╝ ļ░®ļ▓ĢņØä ņé¼ņÜ®ĒĢśņŚ¼ NKņäĖĒżļź╝ ļ░░ņ¢æĒĢśļŖö ļŗ©ņł£ĒĢ£ ļ░®ļ▓ĢņØä ĻĄ¼ņČĢĒĢśĻ│Āņ×É ĒĢśņśĆļŗż(Fig 1A).
Fig.┬Ā1.
Natural killer cells were obtained through various isolation methods, and natural killer cell therapeutic candidates were identified through culture
A. The schema of NK cell culture, B. Flow cytometry results of various isolation methods for NK cell culture, C. Yield of various NK cell isolation methods. D. Total number of NK cell-negative selected cells and CD3+ depleted cells cultured with NKsol TM
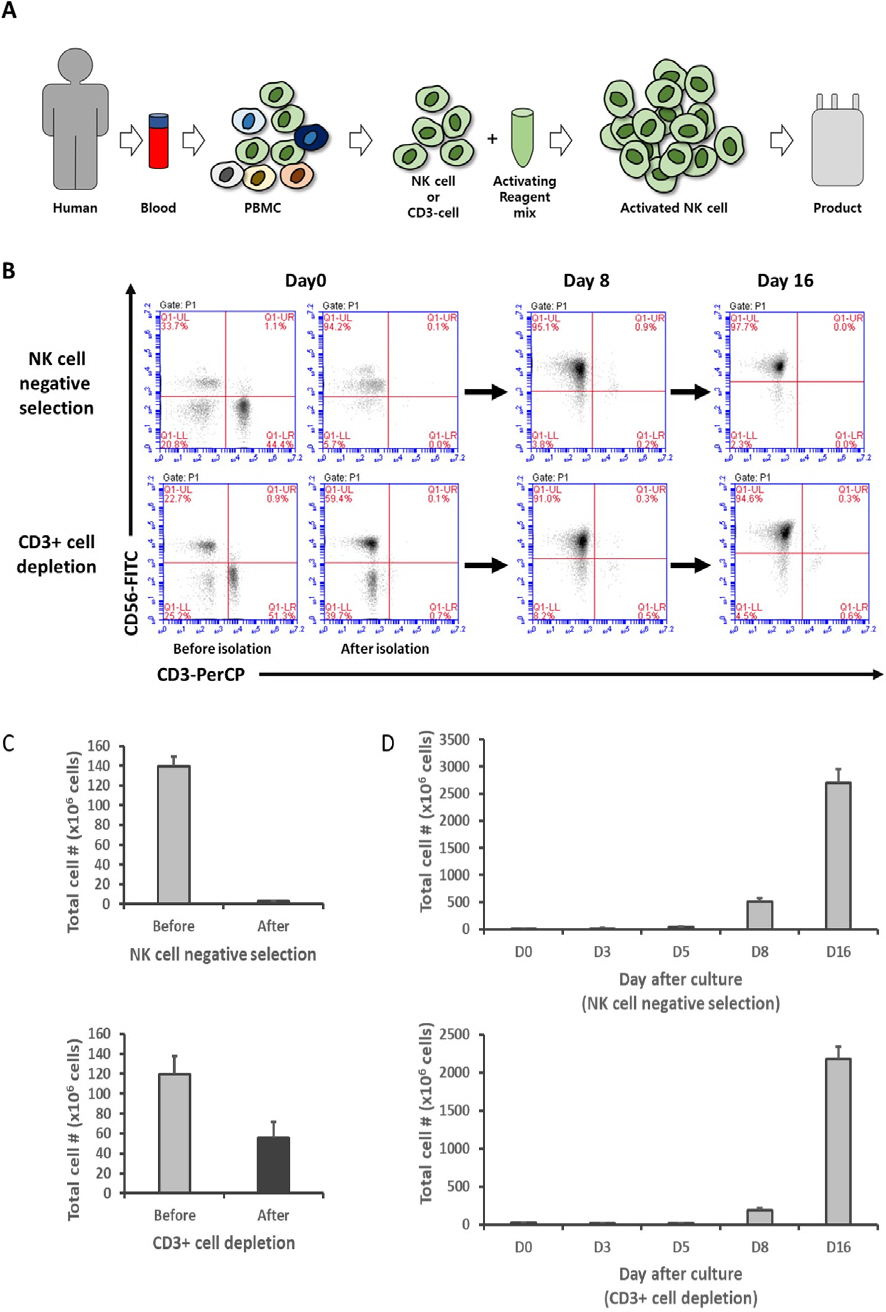
Ficoll-Hypaque ļ░ĆļÅäĻĄ¼ļ░░ ņøÉņŗ¼ļČäļ”¼ļĪ£ ņ¢╗ņØĆ ļ”╝ĒöäĻĄ¼ļź╝ NK cell-negative selection kitņÖĆ CD3 cell positive-selection kitļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņł£ņłśĒĢ£ NKņäĖĒżļź╝ ļ░░ņ¢æĒĢĀ Ēøäļ│┤ ņäĖĒżļōżņØä ļČäļ”¼ĒĢśņśĆļŗż. NK cell-negative selection kitļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ļČäļ”¼ļÉ£ NKņäĖĒżļŖö ļ░░ņ¢æ ņĀä┬ĘĒøä CD56+NKņäĖĒżņØś ļ╣äņ£©ņØ┤ 33.7 %ņŚÉņä£ 94.2 %ļĪ£ ņ”ØĻ░ĆĒ¢łņ£╝ļ®░ ļīĆļČĆļČäņØś CD3+ņäĖĒżņÖĆ CD3-CD56-ņäĖĒżĻ░Ć ņĀ£Ļ▒░ļÉśņ¢┤ ņł£ļÅäĻ░Ć Ēü¼Ļ▓ī ņ”ØĻ░ĆĒĢśņśĆļŗż. ļīĆļČĆļČäņØś CD3+ņäĖĒżļŖö CD3+cell positive selection kitņŚÉ ņØśĒĢ┤ņä£ ņĀ£Ļ▒░ļÉśņŚłļŗż(Fig. 1B). NK cell-negative selectionņŚÉ ņØśĒĢ┤ ļČäļ”¼ļÉ£ ņäĖĒżļŖö ņĢĮ 2 %ņØś ņłśņ£©ņØä ļ│┤ņśĆĻ│Ā, CD3+depletionņØä ĒåĄĒĢ┤ ņ¢╗ņØĆ ņäĖĒżļŖö ņĢĮ 50 %ņØś ņłśņ£©ņØä ļ│┤ņśĆļŗż(Fig. 1C). ņØ┤ļĀćĻ▓ī ļČäļ”¼ļÉ£ Ļ░ü NKņäĖĒż Ēøäļ│┤ĻĄ░ņØä NKņäĖĒż ĒÖ£ņä▒ĒÖö/ņ”ØņŗØ ņŗ£ņĢĮņØĖ NKsol TMņ£╝ļĪ£ ņ▓śļ”¼ĒĢśņŚ¼ ļ░░ņ¢æĒĢ£ Ļ▓░Ļ│╝ NKiņäĖĒżņÖĆ CD3-ņäĖĒż ļ¬©ļæÉ CD56+ NKņäĖĒżļĪ£ ņ£Āņ¦Ć ļ░Å ļČäĒÖöļÉśļŖö Ļ▓āņØä ĒÖĢņØĖĒĢśņśĆļŗż(Fig 1B). NKi ļ░Å CD3-ņäĖĒż ļ¬©ļæÉ ļ░░ņ¢æ Ēøä 16ņØ╝(D16)ņŚÉņä£ ņāüļŗ╣Ē׳ ņ”ØĻ░ĆĒĢ£ ņäĖĒżņ”ØņŗØņØä ļ│┤ņśĆļŗż. ĒŖ╣Ē׳ NiņäĖĒżņØś ņłśļŖö D0ņŚÉņä£ 10├Ś106 ņäĖĒżņśĆņ£╝ļ®░ D16ņŚÉņä£ 2,703.3├Ś106 ņäĖĒżļĪ£ 270ļ░░ ņ”ØĻ░ĆĒĢśņśĆļŗż(Fig. 1D). ļö░ļØ╝ņä£ ņØ┤ Ļ▓░Ļ│╝ļź╝ ĒåĄĒĢ┤ NKņäĖĒż ļ░░ņ¢æņØä ņ£äĒĢ£ Ēøäļ│┤ĻĄ░ņØä ņäĀņĀĢĒĢśĻ│Ā Ļ░äļŗ©ĒĢ£ ļ░░ņ¢æĻ│╝ņĀĢņØä ĒÖĢļ”ĮĒĢśņśĆļŗż.
3.2 Antiviral-related receptor expression comparison of cultured NK cells after NK cell selection and CD3+cell depletion
ņäĀņ▓£ ļ®┤ņŚŁ ņäĖĒżņØĖ NKņäĖĒżļŖö ĻĖēņä▒ SARS-CoV-2Ļ░ÉņŚ╝ņŚÉ ļīĆĒĢ£ ņ┤łĻĖ░ ļ░śņØæņ×ÉļĪ£ņä£ ĒśłņĢĪņŚÉņä£ ĒÅÉļĪ£ ņł£ĒÖśĒĢśļŖö NKņäĖĒżļź╝ ļ¬©ņ¦æĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż. ņØ┤ļ¤¼ĒĢ£ NKņäĖĒżņØś ņØ┤ļÅÖņØĆ ļŗżņ¢æĒĢ£ ņ╝Ćļ¬©ņ╣┤ņØĖņØä ĒåĄĒĢ£ ĒÖöĒĢÖņ£ĀņØĖ(chemoattraction)ņŚÉ ņØśĒĢ┤ ņØ╝ņ¢┤ļé£ļŗż. NKņäĖĒżļŖö ļŗżņ¢æĒĢ£ ĒÖ£ņä▒ĒÖö/ņ¢ĄņĀ£ ņłśņÜ®ņ▓┤ļź╝ ĒåĄĒĢ┤ ļ╣äņĀĢņāü ņäĖĒżļź╝ ņé┤ņāüĒĢśļŖö ļŖźļĀźņØä Ļ░Ćņ¦ĆĻ│Ā ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż.
SARS-CoV-2ņŚÉ Ļ░ÉņŚ╝ļÉ£ ņäĖĒżļŖö HLA-E ļ░£ĒśäņØ┤ ņ”ØĻ░ĆĒĢśļ®░ NKG2CņÖĆ Ļ░ÖņØĆ ņłśņÜ®ņ▓┤ņŚÉ ņØśĒĢ┤ NKņäĖĒżļź╝ ĒÖ£ņä▒ĒÖöĒĢ£ļŗż[10]. SARS-CoV-2 peptideļŖö ļśÉĒĢ£ NKG2D ņłśņÜ®ņ▓┤ņÖĆ Ļ▓░ĒĢ®ĒĢśņŚ¼ NKņäĖĒżņØś ĒÖ£ņä▒ņØä ņ”ØĻ░Ćņŗ£ĒéżļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉśņ¢┤ ņ׳ļŗż[20]. NKG2DņŚÉ ņØśĒĢ┤ ĒÖ£ņä▒ĒÖöļÉ£ NKņäĖĒżļź╝ ņ”ØĒÅŁņŗ£Ēé¼ ņłś ņ׳ļŖö DNAX ļ│┤ņĪ░ ļČäņ×É(DNAM-1), 2B4ļź╝ ĒżĒĢ©ĒĢ£ ņŚ¼ļ¤¼ ĒÖ£ņä▒ĒÖö Ļ│ĄļÅÖ ņłśņÜ®ņ▓┤Ļ░Ć ļ░£ĒśäļÉśĻ│Ā ņ׳ņ£╝ļ®░, ļ░öņØ┤ļ¤¼ņŖż ņĀ£Ļ▒░Ļ░Ć ļŖÉļ”░ ĒÖśņ×ÉņØś NKņäĖĒżļŖö DNAM-1ņØś ļ░£ĒśäĻ│╝ Ļ│ĄļÅÖ ņ¢ĄņĀ£ ņłśņÜ®ņ▓┤ TIGITņØś ļ░£ĒśäņØ┤ Ļ░ÉņåīļÉśņ¢┤ ņ׳ļŗżļŖö ņŚ░ĻĄ¼ Ļ▓░Ļ│╝Ļ░Ć ņ׳ļŗż[21]. ļśÉĒĢ£, NKņäĖĒżļŖö ĒĢŁņ▓┤ ņØśņĪ┤ņä▒ ņäĖĒż ļÅģņä▒ņØä ņ£ĀļÅäĒĢśļŖö Fc-╬│ ņłśņÜ®ņ▓┤ IIIAņØĖ CD16Ļ│╝ ĒĢŁņ▓┤ ņśĄņåīļŗīĒÖö Ēæ£ņĀü ņäĖĒżņŚÉ Ļ▓░ĒĢ®ĒĢśņŚ¼ ĒÖ£ņä▒ĒÖöļÉ£ļŗż. ĒŖ╣Ē׳, CD16ņØĆ ļŗżļźĖ ņłśņÜ®ņ▓┤ļź╝ ĒåĄĒĢ£ ņČöĻ░ĆņĀüņØĖ ĒÖ£ņä▒ĒÖö ņŚåņØ┤ ņŖżņŖżļĪ£ NKņäĖĒżļź╝ ĒÖ£ņä▒ĒÖöĒĢĀ ņłś ņ׳ļŖö ņ£ĀņØ╝ĒĢ£ ņłśņÜ®ņ▓┤ņØ┤ļŗż[22].
NKņäĖĒżļŖö MHC ļÅģļ”ĮņĀüņØĖ ļ¦żņ╗żļŗłņ”śņØä ĒåĄĒĢ┤ ļ░öņØ┤ļ¤¼ņŖż ļŗ©ļ░▒ņ¦ł ĒĢŁņøÉņØä ņØĖņŗØĒĢśĻ│Ā NCR Ļ│äņŚ┤ ņłśņÜ®ņ▓┤(NKp30, NKp44, NKp46)ļŖö ļ░öņØ┤ļ¤¼ņŖżņØś ĻĖĆļ”¼ņ╣Ė ĻĄ¼ņĪ░ņÖĆ ĒĢŁņøÉņØä ņØĖņŗØĒĢśņŚ¼ ļ╣äņĀĢņāü ņäĖĒżņØś ņé¼ļ®ĖņØä ņ£ĀļÅäĒĢśņŚ¼ ļ░öņØ┤ļ¤¼ņŖżņŚÉ Ļ░ÉņŚ╝ļÉ£ ņäĖĒżļź╝ ņĀ£Ļ▒░ĒĢĀ ņłś ņ׳ļŗż[23]. ļśÉĒĢ£, Ēæ£ņĀü ņäĖĒżņØś ņäĖĒż ņÜ®ĒĢ┤ļź╝ ļ¦żĻ░£ĒĢśļŖö Fas ļ”¼Ļ░äļō£(FasL)ņÖĆ ņóģņ¢æ Ļ┤┤ņé¼ ņØĖņ×É Ļ┤ĆļĀ© ņäĖĒżņé¼ļ®Ė ņ£ĀļÅä ļ”¼Ļ░äļō£(TRAIL)ļź╝ ĒżĒĢ©ĒĢ£ ņŚ¼ļ¤¼ ņäĖĒżņÖĖ ļ”¼Ļ░äļō£ļź╝ ļ░£ĒśäĒĢśņŚ¼ ļ░öņØ┤ļ¤¼ņŖżņŚÉ Ļ░ÉņŚ╝ļÉśņ¢┤ ņé¼ļ®Ė ņłśņÜ®ņ▓┤ļź╝ ļ░£ĒśäĒĢśļŖö Ļ░ÉņŚ╝ļÉ£ ņäĖĒżņØś ņé¼ļ®ĖņØä ņ£ĀļÅäĒĢ£ļŗż[24].
CD57ņØĆ ņ”ØņŗØ ļŖźļĀźņØ┤ ņóŗņ¦Ć ņĢŖņØĆ ļģĖĒÖöļÉ£ ņäĖĒż ņāüĒā£ļź╝ ļéśĒāĆļé╝ ņłś ņ׳ļŖö ņ¦ĆĒæ£ļĪ£ ņé¼ņÜ®ļÉśĻ│Ā ņ׳ņ£╝ļ®░, NKņäĖĒżņØś ĻĖ░ļŖźņĀü ļ░śņØæņØä ĒśĢņä▒ĒĢśļŖö ņŻ╝ņÜö ņłśņÜ®ņ▓┤ņØĖ NKG2AņØś ļ░£ĒśäĻ│╝ ļ░śļ╣äļĪĆĒĢ£ļŗżĻ│Ā ļ│┤Ļ│ĀļÉśņŚłļŗż[25].
NKi ļ░Å CD3-ņäĖĒżņØś ĒĢŁļ░öņØ┤ļ¤¼ņŖż Ļ┤ĆļĀ© ņłśņÜ®ņ▓┤ļź╝ ņ£ĀņäĖĒżļČäņäØĻĖ░ļĪ£ ļČäņäØĒĢśņśĆļŗż. ļČäņäØ Ļ▓░Ļ│╝ ĒĢŁļ░öņØ┤ļ¤¼ņŖż ĒÜ©Ļ│╝ļź╝ ļéśĒāĆļé┤ļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņ¦ä ņłśņÜ®ņ▓┤ņØś ļ░£ĒśäņØ┤ PBMC (ļīĆņĪ░ĻĄ░)ņŚÉ ļ╣äĒĢ┤ ņ£ĀņØśĒĢśĻ▓ī ņ”ØĻ░ĆĒĢśļŖö Ļ▓āņØä ĒÖĢņØĖĒĢśņśĆļŗż (Fig 2A, B). NKi ļ░Å CD3-ņäĖĒżļ│┤ļŗż PBMCņŚÉņä£ņØś ļåÆņØĆ CD57 ļ░£ĒśäņØĆ ļÅÖņØ╝ĒĢ£ ļ░░ņ¢æ ĻĖ░Ļ░äņŚÉņä£ ļ╣äĻĄÉĒĢ┤ ļ│╝ ļĢī ļģĖĒÖöĻ░Ć ļŹö ņ¦äĒ¢ēļÉśņŚłņØīņØä ļéśĒāĆļéĖļŗż. ņØ┤ļ¤¼ĒĢ£ Ļ▓░Ļ│╝ļŖö NKi ļ░Å CD3-ņäĖĒżĻ░Ć ĒĢŁļ░öņØ┤ļ¤¼ņŖż ĻĖ░ļŖźņØä Ļ░Ćņ¦ł ņłś ņ׳ņØīņØä ņŗ£ņé¼ĒĢ£ļŗż.
Fig.┬Ā2.
Comparison of antiviral receptor expression in NKi and CD3-cells.
Figures A and B. NKi and CD3-cells gated on CD56- FITC-positive cells and reached with PE-conjugated antibody corresponding to each receptor and PE-positive cells were analyzed by flowcytometry.
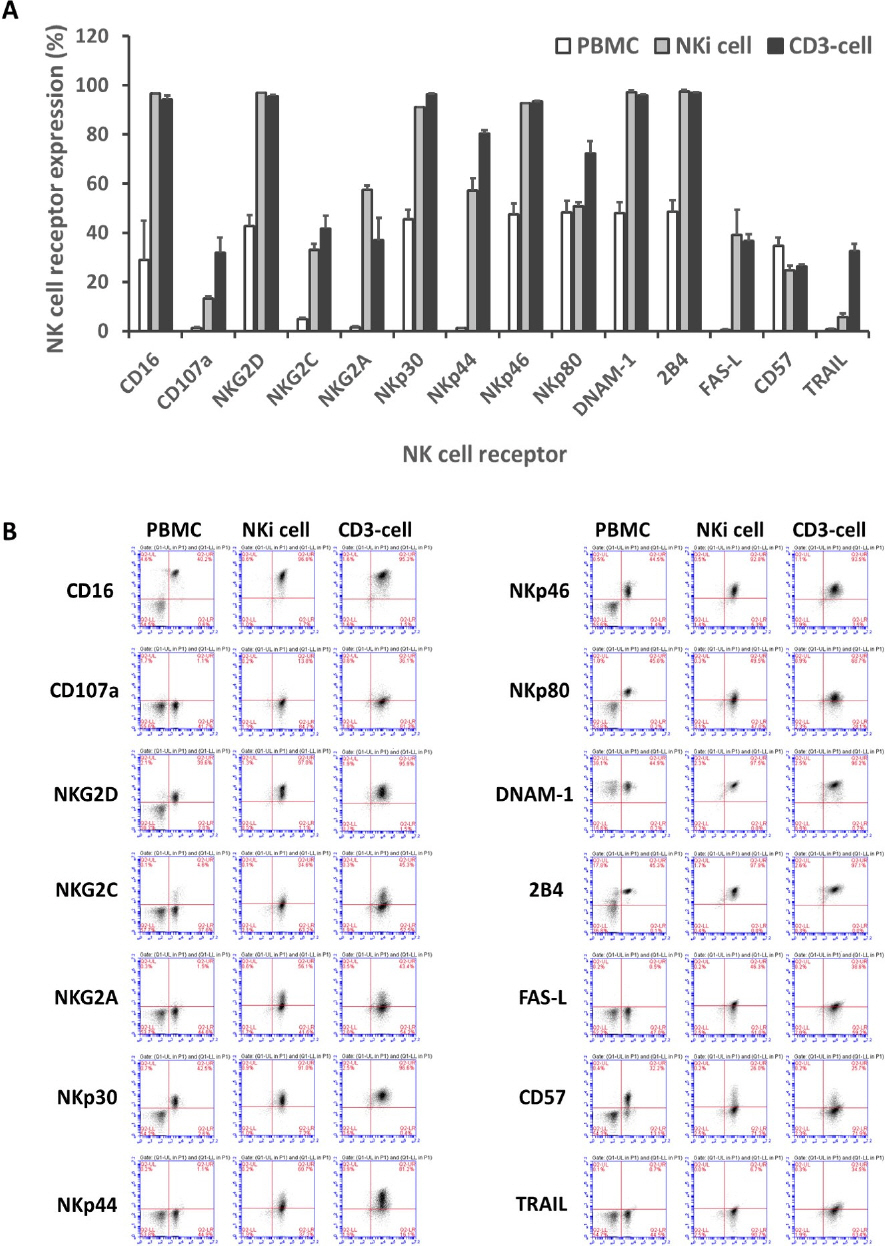
ļö░ļØ╝ņä£ ņØ┤ļōż ņäĖĒżņØś ĒĢŁļ░öņØ┤ļ¤¼ņŖż ĒÜ©ļŖźņØä SARS-CoV-2ņŚÉ Ļ░ÉņŚ╝ļÉ£ ņäĖĒżņÖĆ ļ¦łņÜ░ņŖż ļ¬©ļŹĖņØä ĒåĄĒĢ┤ ĒÖĢņØĖĒĢśņśĆļŗż.
3.3 Antiviral effect of natural killer cell candidate
NKi ļ░Å CD3-ņäĖĒżņØś ĒĢŁļ░öņØ┤ļ¤¼ņŖż ĒÜ©Ļ│╝ļź╝ ĒÖĢņØĖĒĢśĻĖ░ ņ£äĒĢ┤ TCID50 ļČäņäØņØä ņłśĒ¢ēĒĢśņśĆļŗż. VERO E6ņäĖĒżļŖö SARS-CoV-2 ļ░öņØ┤ļ¤¼ņŖżņØś Ēæ£ņĀüņØĖ angiotension-converting enzyme 2(ACE2)ļź╝ ļ░£ĒśäĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż[26]. 100 % confluenceļĪ£ ļ░░ņ¢æļÉ£ VERO E6ņäĖĒżļŖö 1├Ś104 PFU/mLņØś SARS-CoV-2ļĪ£ Ļ░ÉņŚ╝ņŗ£ņ╝░ļŗż. NKi ļ░Å CD3-ņäĖĒżļŖö ņØ┤ĒÄÖĒä░ņäĖĒż(E)ļĪ£ Ēæ£ņĀüņäĖĒż(T)ņØĖ SARS-CoV-2ņŚÉ Ļ░ÉņŚ╝ļÉ£ VERO E6ņÖĆ ĒĢ©Ļ╗ś E:T ļ╣äņ£©ņØ┤ 1:1, 5:1ņØ┤ ļÉśļÅäļĪØ Ļ│Ąļ░░ņ¢æ ņŗ£ņ╝░ļŗż. Ļ│Ąļ░░ņ¢æ Ēøä ĒÜŹļōØĒĢ£ viral supernatant ļŖö ņāłļĪ£ņÜ┤ VERO E6 ņäĖĒżņŚÉ ņ×¼Ļ░ÉņŚ╝ņŗ£ĒéżĻ│Ā, virus titer ņ░©ņØ┤ļź╝ TCID50 ļČäņäØņØä ĒåĄĒĢ┤ ĒÖĢņØĖĒĢśņśĆļŗż. Virus titerļŖö NKi ļ░Å CD3-ņäĖĒżļź╝ Ļ│ĄļÅÖ ļ░░ņ¢æĒĢ©ņ£╝ļĪ£ņŹ© Ēü¼Ļ▓ī Ļ░ÉņåīĒĢ£ Ļ▓āņØä ĒÖĢņØĖĒĢśņśĆļŗż. NKiņäĖĒżņØś Ļ▓ĮņÜ░ logTCID50 Ļ░ÆņØ┤ ļīĆņĪ░ĻĄ░ņŚÉ ļ╣äĒĢ┤ E:T ļ╣äņ£© 1:1Ļ│╝ 5:1ņŚÉņä£ Ļ░üĻ░ü 7.60ņŚÉņä£ 6.67, 5.90ņ£╝ļĪ£ Ļ░ÉņåīĒĢśņśĆļŗż. ņ”ē, SARS-CoV-2 virus titer ļŖö NKiņäĖĒżņŚÉ ņØśĒĢ┤ E:Tļ╣äņ£© 5:1ņŚÉņä£ ņĢĮ 60ļ░░ Ļ░ÉņåīĒĢ£ ņłśņ╣śļź╝ ļ│┤ņśĆļŗż. ļśÉĒĢ£ CD3-ņäĖĒżņØś Ļ▓ĮņÜ░ logTCID50 Ļ░ÆņØ┤ E:T ļ╣äņ£© 1:1Ļ│╝ 5:1ņŚÉņä£ Ļ░üĻ░ü 7.60ņŚÉņä£ 6.66, 6.84ļĪ£ Ļ░ÉņåīĒĢśņśĆļŗż. ņ”ē, SARS-CoV-2 virus titerļŖö CD3-ņäĖĒżņŚÉ ņØśĒĢ┤ ņĢĮ 10ļ░░ Ļ░ÉņåīĒĢ£ ņłśņ╣śļź╝ ļ│┤ņśĆļŗż(Fig. 3A).
Fig.┬Ā3.
Antiviral effect of natural killer cell candidates
A. Viral titers were analyzed in supernatant obtained by co-culture of control, NKi and CD3- cells with SARS-CoV-2 infected VERO E6 cells using the TCID50 assay. B. To confirm the SARS-CoV-2 in vivo treatment effect of NK cell candidates, ACE-tg mice were infected with the virus and survival was observed for two weeks. C. Weight changes and survival were observed following the infusion of NK cell candidates into ACE-tg mice infected with SARS-CoV-2. Control, PBMCs; NKi cells, selected NK cells; CD3-cells, CD3 depleted cells. ***p<0.001.
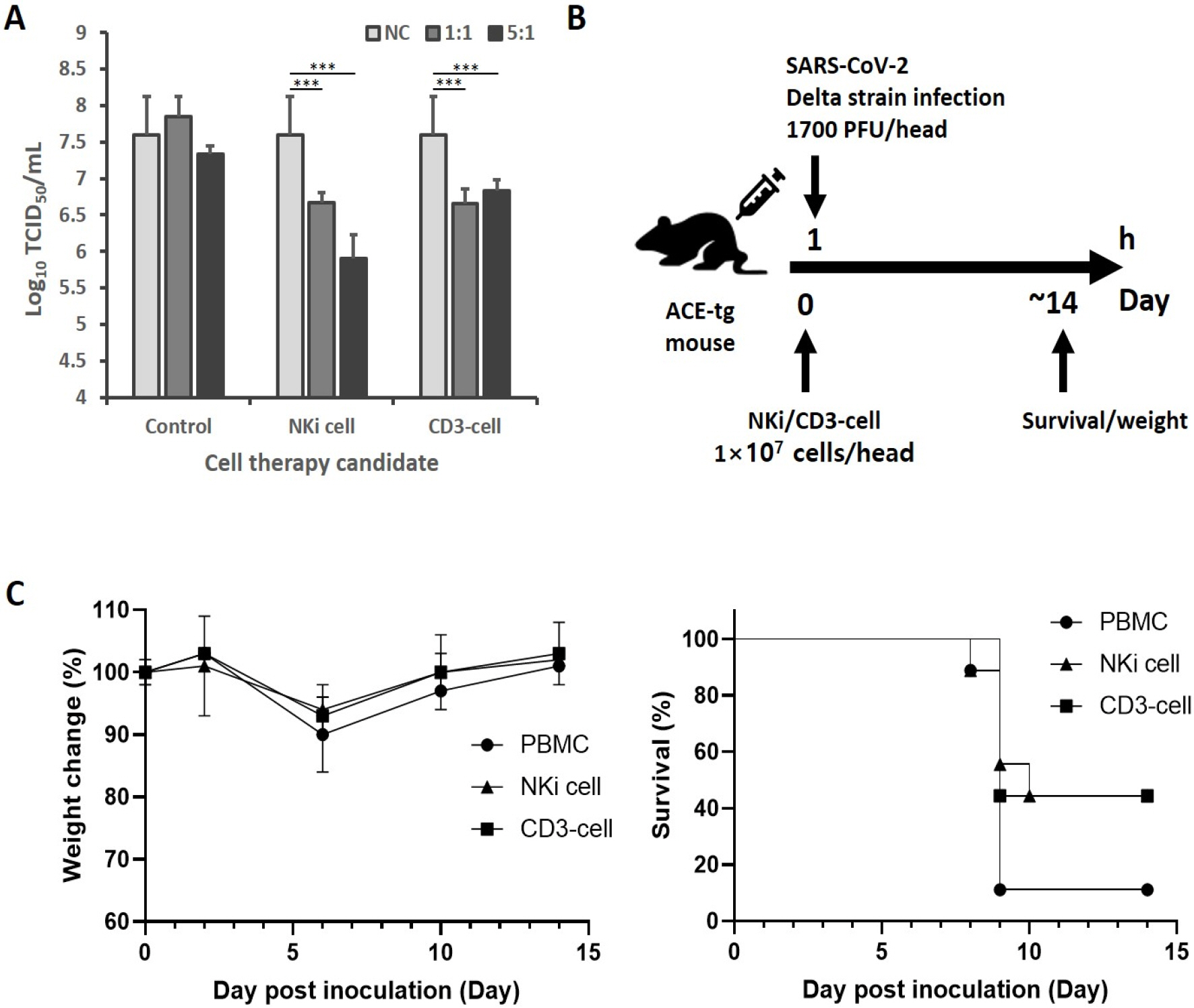
NKiņäĖĒżņÖĆ CD3-ņäĖĒżņØś in vivo ĒĢŁļ░öņØ┤ļ¤¼ņŖż ĒÜ©Ļ│╝ļŖö SARS-CoV-2 ļ░öņØ┤ļ¤¼ņŖż delta variantņŚÉ Ļ░ÉņŚ╝ļÉ£ hACE-transgenic(ACE-tg) ļ¦łņÜ░ņŖż ļ¬©ļŹĖņØä ņé¼ņÜ®ĒĢśņŚ¼ PBMC(ļīĆņĪ░ĻĄ░) ĻĘĖļŻ╣Ļ│╝ ļ╣äĻĄÉĒĢśņŚ¼ ĒÖĢņØĖĒĢśņśĆļŗż. ņØ┤ ņŗżĒŚśņŚÉņä£ļŖö SARS-CoV-2ņŚÉ Ļ░ÉņŚ╝ļÉ£ ACE-tg ļ¦łņÜ░ņŖżņŚÉņä£ NKņäĖĒż Ēøäļ│┤ļ¼╝ņ¦łĻ│╝ PBMCļź╝ 1ĒÜī Ēł¼ņŚ¼ĒĢśĻ│Ā ņ▓┤ņżæ ļ│ĆĒÖöņÖĆ ņāØņĪ┤ņ£©ņØä 2ņŻ╝ ļÅÖņĢł ĻĖ░ļĪØĒĢśņśĆļŗż(Fig. 3B). NKi ļ░Å CD3-ņäĖĒżĻĄ░ ļ¬©ļæÉ ļ¦łņÜ░ņŖżņØś ņ▓┤ņżæņØĆ PBMCĻĄ░Ļ│╝ ņ£ĀņØśļ»ĖĒĢ£ ņ░©ņØ┤Ļ░Ć ņŚåņŚłņ¦Ćļ¦ī, ņāØņĪ┤ņ£©ņØĆ PBMCĻĄ░ņŚÉ ļ╣äĒĢ┤ ļæÉ Ēøäļ│┤ĻĄ░ ļ¬©ļæÉņŚÉņä£ ņ£ĀņØśĒĢśĻ▓ī ņ”ØĻ░ĆĒĢśņśĆļŗż. SARS-CoV-2 ļ░öņØ┤ļ¤¼ņŖż Ļ░ÉņŚ╝ 8ņØ╝ Ēøä ņŻĮņØĆ Ļ░£ņ▓┤Ļ░Ć ņ▓śņØī Ļ┤Ćņ░░ļÉśņŚłņ£╝ļ®░, ļīĆņĪ░ĻĄ░ PBMC ĻĘĖļŻ╣ņØś ņĄ£ņóģ ņāØņĪ┤ļźĀ 11 %ņŚÉ ļ╣äĒĢ┤ņä£ NKi ļ░Å CD3-ņäĖĒż ĻĘĖļŻ╣ņØĆ 44 %ļĪ£ ņāØņĪ┤ņ£©ņØ┤ ļåÆņĢśļŗż(Fig. 3C). ņØ┤ļź╝ ĒåĄĒĢ┤ SARS-CoV-2 Ļ░ÉņŚ╝ ļ¦łņÜ░ņŖżņØś ņāØņĪ┤ņ£©ņØ┤ NKņäĖĒż 1ĒÜī Ēł¼ņŚ¼ļ¦īņ£╝ļĪ£ļÅä Ēü¼Ļ▓ī ņ”ØĻ░ĆĒĢ©ņØä ĒÖĢņØĖĒĢśņśĆļŗż.
4. Ļ▓░ ļĪĀ
COVID-19 ĒÖĢņé░ņ£╝ļĪ£ ņ×ÉĻĖ░ ļ®┤ņŚŁņØ┤ ņżæņÜöĒĢśĻ▓ī ņØĖņŗØļÉśļ®┤ņä£ ņ▓┤ļé┤ņŚÉ ņĪ┤ņ×¼ĒĢśļŖö ļ®┤ņŚŁ ņäĖĒżĻ░Ć ņ×¼ņĪ░ļ¬ģņØä ļ░øĻ│Ā ņ׳ļŗż. ĒŖ╣ļ│äĒĢ£ ņ╣śļŻī ņĢĮ ņŚåņØ┤ļÅä ļ®┤ņŚŁļĀźņ£╝ļĪ£ ņØ┤Ļ▓©ļéĖ Ļ▓ĮņÜ░ļź╝ ņé┤ĒÄ┤ļ│┤ļ®┤, ļ®┤ņŚŁ ņŗ£ņŖżĒģ£ņØ┤ ņĀĢņāüņĀüņ£╝ļĪ£ ņ×æļÅÖĒĢśļ®┤ Ļ▓Įņ”Ø ļé┤ņ¦ĆļŖö ļ¼┤ņ”Øņāüņ£╝ļĪ£ ņÖäņ╣śļÉĀ ņłś ņ׳ĻĖ░ ļĢīļ¼ĖņØ┤ļŗż. ļ®┤ņŚŁņäĖĒżļŖö ņóģņ¢æņØ┤ļéś ļ░öņØ┤ļ¤¼ņŖżņŚÉ Ļ░ÉņŚ╝ļÉ£ ļ╣äņĀĢņāü ņäĖĒżļź╝ ņŻĮņØ┤ļŖö ņŚŁĒĢĀņØä ĒĢśļ®░, ņé¼ņØ┤ĒåĀņ╣┤ņØĖņ£╝ļĪ£ ņ▓┤ļé┤ ļ®┤ņŚŁļĀźņØä ņś¼ļĀżņŻ╝Ļ▒░ļéś ļŗżņ¢æĒĢ£ ņןĻĖ░ņÖĆ ņĪ░ņ¦üņŚÉņä£ ĻĖ░ļŖźĒĢśļŖö ņäĖĒżņØ┤ļŗż. ĒŖ╣Ē׳ ļ®┤ņŚŁņäĖĒż ņżæ NKņäĖĒżļŖö ĒāĆĻ│Āļé£ ļ®┤ņŚŁļ░śņØæņØś ĻĘ╝Ļ░äņ£╝ļĪ£ ļ¬ĖņØś ļ®┤ņŚŁĻ│äņŚÉņä£ ļ░®ņ¢┤ 1ņäĀņØä ļŗ┤ļŗ╣ĒĢśĻ│Ā ņ׳ļŖö ņäĖĒżļĪ£ ņóģņ¢æņäĖĒżļéś ļ░öņØ┤ļ¤¼ņŖż Ļ░ÉņŚ╝ ņäĖĒż ļō▒ ļ╣äņĀĢņāü ņäĖĒżļź╝ ņ”ēĻ░üņĀüņ£╝ļĪ£ ņØĖņŗØĒĢ┤ ņĀ£Ļ▒░ĒĢ£ļŗż[10,27].
ļö░ļØ╝ņä£ ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö NK cell-negative selectionĻ│╝ CD3 cell-positive selectionņØä ņØ┤ņÜ®ĒĢśņŚ¼ NKņäĖĒż ļ░░ņ¢æ Ēøäļ│┤ĻĄ░ņØä ņäĀņĀĢĒĢśĻ│Ā ņ”ØņŗØ/ĒÖ£ņä▒ĒÖö ņŗ£ņ╝£ ņäĖĒżņ”ØņŗØ, ĒĢŁļ░öņØ┤ļ¤¼ņŖż ņłśņÜ®ņ▓┤ ļ░£ĒśäņØä ļČäņäØĒĢśĻ│Ā SARS-CoV-2 Ļ░ÉņŚ╝ņŚÉ ļīĆĒĢ£ ĒĢŁļ░öņØ┤ļ¤¼ņŖż ĒÜ©ļŖźņØä in vitro ĻĘĖļ”¼Ļ│Ā in vivoņŚÉņä£ ĒÖĢņØĖĒĢśņŚ¼ ĒĢŁļ░öņØ┤ļ¤¼ņŖż ņäĖĒżņ╣śļŻīņĀ£ Ēøäļ│┤ĻĄ░ņØś ņ£ĀĒÜ©ņä▒ņØä ĒÖĢņØĖĒĢśņśĆļŗż.
ņäĖĒż ņ×ģņ¢æļ®┤ņŚŁ ņ╣śļŻīņŚÉņä£ļŖö NKņäĖĒżļź╝ ĒÜ©ņ£©ņĀüņ£╝ļĪ£ ņ”ØņŗØņŗ£ņ╝£ ļīĆļ¤ēņ£╝ļĪ£ ļ░░ņ¢æĒĢśļŖö Ļ▓āņØ┤ ņżæņÜöĒĢśļŗż. Ļ▒┤Ļ░ĢĒĢ£ Ļ│ĄņŚ¼ņ×ÉļĪ£ļČĆĒä░ ņ¢╗ņØĆ ĒśłņĢĪņ£╝ļĪ£ļČĆĒä░ NKiņäĖĒżņÖĆ CD3-ņäĖĒżļź╝ ĒÜŹļōØĒĢśņśĆĻ│Ā, ņ”ØņŗØĻ│╝ ĒÖ£ņä▒ĒÖö ļ¼╝ņ¦łņØĖ NKsol TMņØä ņ▓śļ”¼ĒĢśņŚ¼ NKņäĖĒżļź╝ ļ░░ņ¢æĒĢśņśĆļŗż. ņØ┤ļź╝ ĒåĄĒĢ┤ NKi ļ░Å CD3-ņäĖĒżļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ļ¦żņÜ░ ņł£ņłśĒĢ£ NKņäĖĒżļź╝ ļīĆļ¤ēņ£╝ļĪ£ ņ”ØņŗØņŗ£Ēé¼ ņłś ņ׳ņØīņØä ĒÖĢņØĖĒĢśņśĆļŗż(Fig. 1)
NKņäĖĒżļŖö ĒÖ£ņä▒ĒÖö/ņ¢ĄņĀ£ ņłśņÜ®ņ▓┤ņØś ļŗżņ¢æĒĢ£ ļ”¼Ļ░äļō£ ņāüĒśĖņ×æņÜ®ņØä ĒåĄĒĢ┤ ļ╣äņĀĢņāü ņäĖĒżļź╝ ņé┤ņāüĒĢśļŖö ļŖźļĀźņØä Ļ░Ćņ¦ĆĻ│Ā ņ׳ļŗż. SARS-CoV-2ņŚÉ Ļ░ÉņŚ╝ļÉ£ ņäĖĒżļŖö HLA-EņØś ļ░£ĒśäņØ┤ ņ”ØĻ░ĆĒĢśņŚ¼ NKņäĖĒżņØś NKG2Cļź╝ ĒÖ£ņä▒ĒÖö ņŗ£Ēé¼ ņłś ņ׳ļŗż[10]. ļśÉĒĢ£ SARS-CoV-2 peptideļŖö NKņäĖĒżņØś NKG2D ņłśņÜ®ņ▓┤ņŚÉ Ļ▓░ĒĢ®ĒĢśņŚ¼ NKņäĖĒżņØś ĒÖ£ņä▒ņØä ņ”ØĻ░Ćņŗ£Ēé©ļŗżļŖö Ļ▓āņØ┤ ņĢīļĀżņĀĖ ņ׳ļŗż[20]. ļ░öņØ┤ļ¤¼ņŖż ņĀ£Ļ▒░Ļ░Ć ļŖÉļ”░ ĒÖśņ×ÉņŚÉņä£ NKņäĖĒżņØś DNAM-1 Ļ░ÉņåīņÖĆ TIGIT ļ░£ĒśäņØ┤ ņ”ØĻ░ĆĒĢśņŚ¼ ņ׳ļŖö Ļ▓āņØ┤ ļ│┤Ļ│ĀļÉ£ ļ░ö ņ׳ļŗż[21]. NKņäĖĒżļŖö ļśÉĒĢ£ MHC ļÅģļ”ĮņĀüņØĖ ļ®öņ╗żļŗłņ”śņ£╝ļĪ£ ļ░öņØ┤ļ¤¼ņŖż ĒĢŁņøÉņØä ņØĖņŗØĒĢĀ ņłś ņ׳Ļ│Ā NCR Ļ│äņŚ┤ ņłśņÜ®ņ▓┤(NKp30, NKp44, NKp46)ņŚÉ ņØśĒĢ┤ ļ░öņØ┤ļ¤¼ņŖż Ēü┤ļ”¼ņ╣Ė ĻĄ¼ņĪ░ ļ░Å ĒĢŁņøÉņØä ņØĖņŗØĒĢśņŚ¼ ļ░öņØ┤ļ¤¼ņŖż Ļ░ÉņŚ╝ ņäĖĒżļź╝ ņĀ£Ļ▒░ĒĢĀ ņłś ņ׳ļŗż[23]. ņ”ØņŗØ ļ░Å ĒÖ£ņä▒ĒÖöļÉ£ NKiņäĖĒżņÖĆ CD3-ņäĖĒżļŖö PBMCņÖĆ ļ╣äĻĄÉĒĢśņŚ¼ ĒĢŁļ░öņØ┤ļ¤¼ņŖżņÖĆ Ļ┤ĆļĀ©ļÉ£ ĒÖ£ņä▒ĒÖö ņłśņÜ®ņ▓┤ņØś ņāüļŗ╣ĒĢ£ ņ”ØĻ░Ćļź╝ ļ│┤ņśĆļŗż. ļö░ļØ╝ņä£ ņØ┤ Ļ▓░Ļ│╝ļŖö NKiņäĖĒżņÖĆ CD3-ņäĖĒżĻ░Ć ĒĢŁļ░öņØ┤ļ¤¼ņŖż ĒÜ©ļŖźņØä Ļ░Ćņ¦ł ņłś ņ׳ņØīņØä ļ│┤ņŚ¼ņżĆļŗż.
ļ░öņØ┤ļ¤¼ņŖż titerņØś ļ│ĆĒÖöļŖö SARS-CoV-2 Ļ░ÉņŚ╝ņäĖĒżņÖĆ NK ņäĖĒżņ╣śļŻīņĀ£ Ēøäļ│┤ĻĄ░ņØĖ NKiņäĖĒżņÖĆ CD3-ņäĖĒżņØś Ļ│Ąļ░░ņ¢æņØä ĒåĄĒĢ┤ Ļ░Éņåīļź╝ ĒÖĢņØĖĒĢśņśĆļŗż. NKņäĖĒż Ēøäļ│┤ĻĄ░ņØĆ SARS-CoV-2 ļ░öņØ┤ļ¤¼ņŖżĻ░Ć Ļ░ÉņŚ╝ļÉ£ VERO E6 ņäĖĒżļź╝ ņé┤ņāüĒĢśņŚ¼ ļ░öņØ┤ļ¤¼ņŖż titerĻ░Ć Ēü¼Ļ▓ī Ļ░ÉņåīĒĢśņśĆļŗż. NKiņäĖĒżņØś Ļ▓ĮņÜ░ 60ļ░░ ļé«ņØĆ ļ░öņØ┤ļ¤¼ņŖż titerĻ░Ć Ļ░ÉņåīĒĢśņśĆĻ│Ā, CD3-ņäĖĒżņØś Ļ▓ĮņÜ░ ņĢĮ 10ļ░░ ļé«ņØĆ ļ░öņØ┤ļ¤¼ņŖż titer Ļ░Éņåīļź╝ ļ│┤ņśĆļŗż(Fig. 3A). in vivoņŚÉņä£ NKņäĖĒż Ēøäļ│┤ĻĄ░ņØś ļ░öņØ┤ļ¤¼ņŖż ņĀ£Ļ▒░ ĒÜ©Ļ│╝ļź╝ ĒÖĢņØĖĒĢśĻĖ░ ņ£äĒĢ┤ SARS-CoV-2ņŚÉ Ļ░ÉņŚ╝ļÉ£ ACE-tg ļ¦łņÜ░ņŖż ļ¬©ļŹĖņØä ņé¼ņÜ®ĒĢśņŚ¼ ņāØņĪ┤ņ£©ņØä ĒÖĢņØĖĒĢśņśĆļŗż. SARS-CoV-2ņŚÉ Ļ░ÉņŚ╝ļÉ£ ACE-tg ļ¦łņÜ░ņŖżņŚÉ NKi ļ░Å CD3-ņäĖĒżļź╝ ļŗ©ĒÜī Ēł¼ņŚ¼ĒĢ£ Ļ▓░Ļ│╝ ņ▓┤ņżæņØś ļ│ĆĒÖö ņŚåņØ┤ ļīĆņĪ░ĻĄ░ļ│┤ļŗż ļŹö ļåÆņØĆ ņāØņĪ┤ņ£©ņØä ļ│┤ņśĆļŗż(Fig. 3C). ņØ┤ Ļ▓░Ļ│╝ļŖö NKņäĖĒżņØś ļŗ©ĒÜī Ēł¼ņŚ¼ļĪ£ SARS-CoV-2 Ļ░ÉņŚ╝ņŚÉ ļīĆĒĢ£ ņāØņĪ┤ņ£©ņØä ļåÆņØ╝ ņłś ņ׳ņØīņØä ĒÖĢņØĖĒĢ£ Ļ▓āņØ┤ļŗż. ĒĢśņ¦Ćļ¦ī ņČöĻ░ĆņĀüņ£╝ļĪ£ ĒĢŁļ░öņØ┤ļ¤¼ņŖż ĒÜ©ļŖźņØä ļåÆņØ┤ĻĖ░ ņ£äĒĢ┤ņä£ļŖö ļŗżņżæĒł¼ņŚ¼ ļśÉļŖö ņÜ®ļ¤ē ņ”ØĻ░ÉņØä ĒåĄĒĢ┤ ĒÜ©ņ£©ņĀüņØĖ NKņäĖĒżņØś ņÜ®ļ▓ĢņØä ĒÖĢļ”ĮĒĢ┤ņĢ╝ ĒĢĀ ĒĢäņÜöĻ░Ć ņ׳ļŗż.
ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ņÜ░ļ”¼ļŖö ļ¬ć Ļ░Ćņ¦Ć NKņäĖĒżņØś ņäĀļ│ä ļ░®ļ▓ĢņØä ĒåĄĒĢ┤ NKņäĖĒżņØś ĻĖ░ņøÉņØä ņäĀņĀĢĒĢśņŚ¼ ņ”ØņŗØ ļ░Å ĒÖ£ņä▒ĒÖöļź╝ ĒåĄĒĢ┤ ļīĆļ¤ēņØś NKņäĖĒżļź╝ ļ░░ņ¢æĒĢĀ ņłś ņ׳ļŖö Ēøäļ│┤ĻĄ░ņØä ĒÖĢļ”ĮĒĢśņśĆļŗż. NKsol TMņ£╝ļĪ£ ļ░░ņ¢æĒĢ£ NKņäĖĒż Ēøäļ│┤ĻĄ░ņØĖ NKiņäĖĒżņÖĆ CD3-ņäĖĒżļŖö ļåÆņØĆ ņł£ļÅäļĪ£ ņāüļŗ╣Ē׳ ņ”ØĻ░ĆĒĢ£ ĒÖ£ņä▒ĒÖö ņłśņÜ®ņ▓┤ņØś ļ░£ĒśäņØä ļ│┤ņśĆļŗż. in vitro/in vivo Ļ▓░Ļ│╝ SARS-CoV-2 Ļ░ÉņŚ╝ ņäĖĒżļź╝ ņé┤ņāüĒĢśņŚ¼ ļ░öņØ┤ļ¤¼ņŖżĻ░Ć ņ×¼ņāØņé░ļÉśņ¦Ć ņĢŖļÅäļĪØ ņ¢ĄņĀ£ĒĢśņŚ¼ ļ░öņØ┤ļ¤¼ņŖż titerĻ░Ć Ēü¼Ļ▓ī Ļ░ÉņåīĒĢśņśĆĻ│Ā SARS-CoV-2ņŚÉ Ļ░ÉņŚ╝ļÉ£ ļ¦łņÜ░ņŖżņØś ņāØņĪ┤ņ£©ņØä Ēü¼Ļ▓ī ņ”ØĻ░Ćņŗ£ņ╝░ļŗż.
ņÜöņĢĮĒĢśņŚ¼, ņØ┤ ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö ĒĢŁļ░öņØ┤ļ¤¼ņŖż NKņäĖĒżņ╣śļŻīņĀ£ņØś ļŗżņ¢æĒĢ£ Ēøäļ│┤ĻĄ░ņØś ļ░░ņ¢æ Ļ│ĄņĀĢņØä ĒÖĢļ”ĮĒĢśĻ│Ā SARS-CoV-2 ļ░öņØ┤ļ¤¼ņŖż Ļ░ÉņŚ╝ļ│æņŚÉ ļīĆĒĢ£ ĒĢŁļ░öņØ┤ļ¤¼ņŖżņĀ£ļĪ£ņØś Ļ░ĆļŖźņä▒ņØä ĒÖĢņØĖĒĢśņśĆļŗż.




